http://s30662109275.mirtesen.ru/blog/43514857321/8--GENETICH...
Смутные сомнения
Когда читаешь о достижениях генетиков в области происхождения человека, порой аж дух захватывает – так много им уже удалось выяснить всего-то за несколько десятков лет. И картина, выстроенная с их помощью, кажется абсолютно достоверной и даже фундаментальной.
Однако если присмотреться повнимательней к деталям различных публикаций, то начинают возникать смутные сомнения – а так ли все у них «безгрешно» на самом деле?..
Возьмем, к примеру, утверждение о том, что предки всех современных людей произошли от некоей очень малочисленной группы, которая обитала в восточной Африке 160-200 тысяч лет назад.
О том, что вообще африканская версия происхождения различных видов предшественника человека построена лишь на основе мнения Дарвина и на ошибочной трактовке местоположения находок, указывалось ранее. Оказывается, если старательно покопаться в интернете, можно обнаружить сообщения, что генетические исследования указывают в качестве «прародины» сапиенсов не только Африку, но и другие континенты!.. Более того, несмотря на достижения генетиков, среди палеоантропологов еще так и не похоронена окончательно так называемая гипотеза полицентризма – гипотеза, допускающая параллельное развитие человечества в разных центрах. Это указывает на то, что выводы генетиков не столь уж убедительны – даже в среде профессиональных палеоантропологов. Подобных публикаций мало, но их небольшое количество можно вполне объяснить не недостатком аргументов для предлагаемых иных вариантов места происхождения Гомо сапиенс, а всего лишь тем, что ныне большинство генетиков (как указывается в некоторых источниках) придерживается версии африканской «прародины». И тут появляется ощущение простой подгонки под дарвиновскую точку зрения…
А теперь обратим внимание на датировки.
Итак, в 1987 году группа американских ученых во главе с А.Уилсоном на основании исследований мтДНК вычислила время расхождения эволюционных линий различных современных внутривидовых (расовых) групп современного человечества и пришла к сенсационному выводу, что все современное человечество происходит от общей древней африканской популяции, причем от одной женщины-праматери, жившей 140-290 тысяч лет назад. Эта женщина-праматерь тут же получила прозвище «Евы». Потомки «Африканской Евы», как считается, расселились по всем континентам, вытесняя более древних предшественников человека, но не смешиваясь с ними.
Позже эксперименты и расчеты, проведенные Уилсоном, неоднократно повторялись в разных вариантах, при этом имели место значительные колебания предполагаемого времени существования этой «Африканской Евы» – по разным исследованиям, 166-249 тысяч лет, 176-222 тысячи лет, 188-270 тысяч лет назад. А результаты некоторых генетических исследований уводят начало линии сапиенса вообще на уровень «гейдельбергского» человека. Так, сотрудник Центра молекулярной генетики Калифорнийского университета в Сан-Диего К.Уилс, применив свой метод оценки древности человеческих популяций, получил для возраста единого очага сапиенсов диапазон от 436 до 806 тысяч лет. Близкое к максимальному значению данного времени образования ветви эволюции в сторону Гомо сапиенс получено другой группой американских исследователей во главе с Дж.М.Фуллертоном – 800 тысяч лет назад.
Поражает размах в различии датировок – не проценты или даже десятки процентов, а в несколько раз!.. Такой разброс результатов выявляет наличие серьезных проблем у самой методики подобных генетических исследований, что бы там ни утверждалось в научных публикациях в ее защиту.
Косвенно данный вывод подтверждает и то, что возраст общего «праотца» всех современных людей – так называемого «Адама», определяемый не по митохондриальной ДНК, передающейся по материнской линии, а по ядерной ДНК (Y-хромосоме, передающейся по мужской линии), серьезно отличается от получаемого в целом по той же методике расчетов возраста «Евы» (хотя есть публикации, в которых указываются и примерно совпадающие даты)…

Рис. 144. Каков же возраст «Адама» и «Евы»?
Более того, в последнее время участились публикации, в которых гипотеза «митохондриальной Евы» подвергается серьезной критике со стороны не только антропологов, но и самих генетиков. Подвергается сомнению и тезис о полном несмешении Гомо сапиенс с неандертальцами и другими обитателями мест, куда приходили сапиенсы.
Так, например, «денисовский» человек хотя и признается самостоятельным подвидом, но имеет свой вклад в геном современного человечества – по некоторым исследованиям до нескольких процентов. А по неандертальцам постепенно начинает доминировать компромиссная точка зрения – некоторые исследователи (например, американский антрополог Л.Брэйс) считают, что генетический вклад неандертальца в Европе мог достигать даже 25 процентов!..
Алан Темплтон из Мичиганского университета еще более категоричен. Он полагает, что по одному-единственному участку генома (например, по мтДНК) нельзя делать окончательные выводы об эволюции и истории расселения человечества. Для таких выводов необходим комплексный анализ многих разных участков генома.
Темплтон утверждает, что ничего удивительного нет ни в «Адаме», ни в «Еве», поскольку любые гомологичные (грубо – схожие) участки ДНК где-нибудь в прошлом неизбежно сходятся в одну точку, то есть в одну предковую молекулу ДНК. И эта точка вовсе не обязательно совпадает с моментом возникновения вида. Более того, если брать разные гомологичные участки ДНК, каждый из них даст свою, отличную от других «точку схождения».
Темплтон показал, что ожидаемое время схождения эволюционного древа, построенного для отдельного участка ДНК, в одну точку, зависит от того, в скольких экземплярах присутствует данный участок в клетках. Быстрее всего должны сходиться как раз мтДНК и Y-хромосома – генетики и наблюдают, что они сходятся примерно 150-160 тысяч лет назад. А, скажем, участки на Х-хромосоме, занимающей промежуточное положение (у женщин она присутствует в двух экземплярах, у мужчин в одном), сходятся в более далеком прошлом – до 2 миллионов лет назад. Есть и участки, которые сходятся в еще более глубокой древности – некоторые даже еще до того, как разделились эволюционные линии человека и шимпанзе.
По результатам исследований Темплтона, три участка ДНК сохранили следы древнейшей волны выхода из Африки около 1,9 миллионов лет назад. Фактически это означает, что в наших жилах течет кровь древних азиатских архантропов. Семь участков ДНК свидетельствуют о втором исходе из Африки около 650 тысяч лет назад – так называемая ашельская экспансия. Представители этой волны – тоже наши прямые предки. Наконец, еще пять участков ДНК (в том числе мтДНК и Y-хромосома) подтверждают третий исход из Африки.
Более того, данные Темплтона показывают, что обмен генами между евразийскими и африканскими популяциями наших предков практически никогда не прекращался, хоть и был сильно затруднен большими расстояниями. Получается, что древнее человечество вовсе не было простой совокупностью отдельных изолированных популяций, а было относительно единым на протяжении двух последних миллионов лет.
Эти выводы Темплтона серьезно противоречат взглядам, господствующим ныне у антропологов…
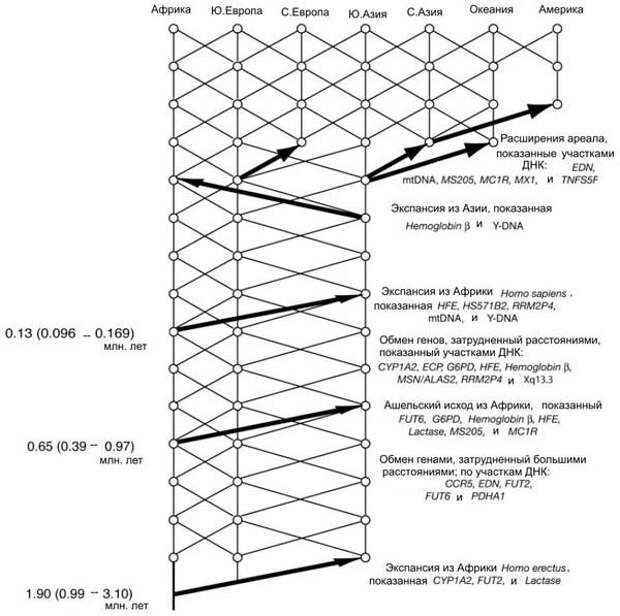
Рис. 145. Схема истории человечества по Темплтону
Из подобной разноголосицы следует одно – современные методы генетических исследований не столь уж однозначны и «безгрешны», как это порой представляется широкой общественности в сообщениях средств массовой информации. Но чтобы понять, в чем именно могут находиться скрытые ошибки и где слабые места этих методов, нам не остается ничего иного, как погрузиться в некоторые их детали.
Перевести «птичий язык» генетиков с его множеством специфических терминов на обычный разговорный язык, доступный рядовому читателю, отнюдь не просто. Но я попробую…
Для этого далее я буду использовать интернетную статью А.Чубенко «Сыновья Адама», где материал изложен в более-менее читаемом виде. Хотя и с ее помощью не удается полностью избежать использования дополнительных терминов.
Некоторые детали генетических методов
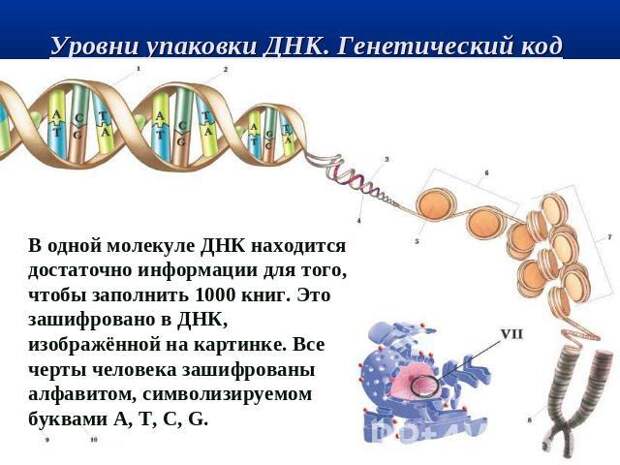
Основной объект генетических исследований – молекулы ДНК (дезоксирибонуклеиновой кислоты). Эти молекулы сосредоточены в клетке двух местах – в хромосомах в ядре (ядерные ДНК – яДНК) и в митохондриях (мтДНК) в цитоплазме клетки.

Рис. 146. Строение клетки человека
Наследственная информация кодируется цепочками двух типов – ДНК и РНК (рибонуклеиновая кислота). Цепочки обоих типов состоят из молекул нуклеотидов (или «азотистых оснований») четырех видов – аденин (A), гуанин (G), цитозин (C) и тимин (T). Таким образом, одинарную цепочку можно представить как произвольную последовательность из четырех букв, например ATCGGATCAAC, полностью отвлекаясь как от химической природы ее элементов, так и от типа химических связей между ними.
Следует отметить, что у любой такой цепи имеется естественная химическая «ориентация», которую вышеприведенная простая запись не отражает. А именно: каждый нуклеотид связан со своими соседями в двух позициях – в месте присоединения фосфата (5-ый атом углеродного цикла) и в месте присоединения радикала OH (там, где находится 3-ий углерод). Если обозначить указанные связи нуклеотидов через 5' и 3' соответственно, то можно просто сформулировать важный факт: соседние нуклеотиды могут соединяться друг с другом только «концами» разных типов, то есть к 3' концу одного может присоединиться 5' конец другого и наоборот, но соединения 5'-5' и 3'-3' невозможны. Используя данные обозначения, говорят о 5'-ом либо 3'-ем конце всей цепи и пишут, например, так: 5'-ATCGGATCAAC-3'. Если условиться, что при буквенной записи цепи начинаются с 5' конца, обозначения 5' и 3' можно опускать. Заметим, что ориентация цепей важна не только как естественный способ их упорядочения, но еще и потому что она учитывается самой природой в процессах передачи генетической информации – новая цепь может строиться только в направлении от 5' к 3'.
Если РНК – одиночная цепь нуклеотидов, то ДНК – это двойная цепочка. Точнее – две цепочки, соединенные друг с другом с соблюдением так называемой комплементарности (дополнительности). Так C есть комплемент (дополнение) к G, G к C, A к T и T к A. Или, говоря другими словами, аденин соединяется только с тимином, гуанин – только с цитозином.
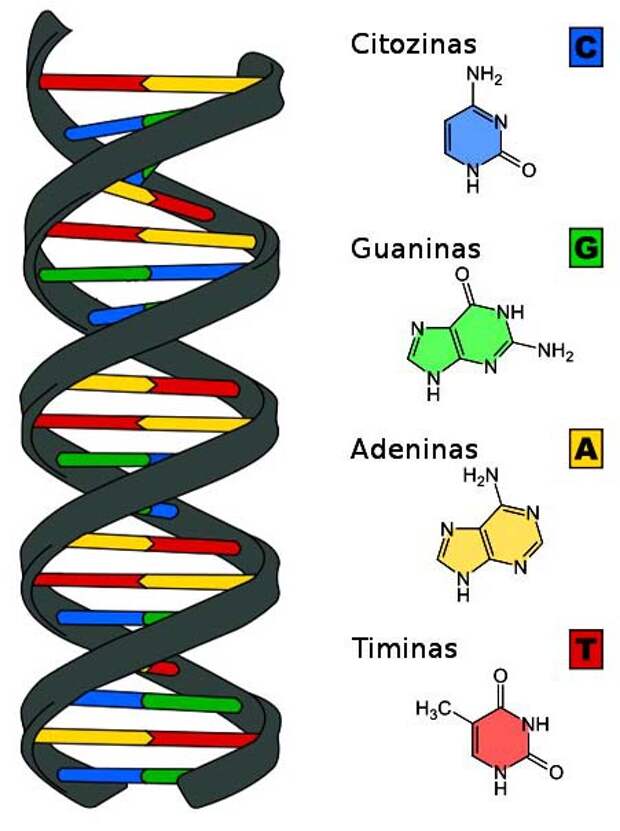
Рис. 147. Нуклеотиды и их соединение в ДНК
При секвенировании (расшифровке) ДНК производится целый ряд весьма непростых процедур, в результате чего появляется длинная запись последовательности расположения нуклеотидов в цепочке – сиквенсы. Эти записи в нескольких форматах ныне хранятся в базах данных сиквенсов, крупнейшей из которых является Генбанк – онлайн-хранилище, доступное исследователям. Ниже приводится пример фрагмента такой записи:
>Homo sapiens mitochondrion, D-loop region, sample #1
AATCAGAGAAAAAGTCTTTAACTCCACCATTAGCACCCAAAGCTAAGATTCTAATTTAAACTATTCTCTGTT
CTTTCATGGGGAAGCAGATTTGGGTACCACCCAAGTATTGACTCACCCATCAACAACCGCTATGTATTTCGT
ACATTACTGCCAGCCACCATGAATATTGTACGGTACCATAAATACTTGACCACCTGTAGTACATAAAAACCC
AATCCACATCAAAATCCCCTCCCCATGCTTACAAGCAGGTACAGCAATCAACCCTCAACTATCACACATCA
ACTGCAACTCCAAAGCCACCCCTCACCCACTAGGATACCAACAAACCTACCCACCCTTAACAGTACATAGT
ACATAAAGCCATTTACCGTACATAGCACATTACAGTCAAATCCCTTCTCGTCCCCATGGATGACCCCC
В ходе эволюционного процесса в живом организме происходят изменения – мутации, которые в дальнейшем передаются по наследству. Наследственная же информация, как указывалось выше, передается с помощью ДНК. Таким образом, в результате мутаций происходит изменение структуры ДНК – изменение последовательности нуклеотидов. Наличие мутаций как раз и выявляется по различию записей (сиквенсов) таких последовательностей, отражающих структуру ДНК у разных людей (животных, растений и т.д.).
«С эволюционной (филогенетической) точки зрения мутациями обычно называют различия между двумя сиквенсами, про которые установлено, что один из них является «непосредственным» эволюционным предком другого. Между ними могут быть десятки и даже сотни трансмиссий, и нередко мы сталкиваемся с ситуацией, когда близкие «родственники» сиквенса не обнаружены, и его приходится сравнивать с достаточно отдаленными, что приводит к реконструированию большого числа мутаций при переходе от узла-предка к его ныне живущим «непосредственным» узлам-потомкам. Поясним сказанное простым примером.
Taxon1 AGCCTGGATCGTATTAGCATGTA
Taxon2 AGCCTGAATCGTACTAGCATGTA
Taxon3 AGCCTGAATCGTATTAGCACGCA…
Разумно предположить, что общий предок трех данных сиквенсов в каждой позиции имел нуклеотид, встречающийся у большинства потомков:
Anc AGCCTGAATCGTATTAGCATGTA
но при этом любой из трех сиквенсов Taxon1, Taxon2 и Taxon3 хотя бы в одной позиции отличается от своего предка. Иными словами, где-то на эволюционном пути от сиквенса Anc к трем его потомкам произошли мутации, а именно, линии Taxon1 и Taxon2 мутировали в одной позиции, а Taxon3 – сразу в двух» (А.Чубенко, «Сыновья Адама»).
Обратим внимание на слова «разумно предположить» в приведенной выше цитате. Спорить не буду – в данном случае это предположение действительно разумно. Однако это – всего лишь предположение, а вовсе не четко установленный факт!.. А любое предположение – даже не просто гипотеза, а субъективная гипотеза!..
Далее мне придется немного перефразировать А.Чубенко, чтобы избежать излишней специфической терминологии.
Усложним задачу и зададимся вопросом – в каком порядке происходили мутации в линии Taxon3? Ответить на данный вопрос, имея в наличии только указанный набор данных, невозможно. Сначала, например, могла произойти мутация в четвертой позиции с конца, а затем во второй с конца, а могло быть и наоборот – сначала во второй с конца, а затем в четвертой, или вообще обе мутации могли произойти одновременно (пусть это и маловероятно).
Однако, если бы в нашем наборе данных присутствовал такой сиквенс:
Taxon4 AGCCTGAATCGTATTAGCATGCA
то можно было бы сделать вывод, что он лежит «на полпути» между вариантом Taxon3 и общим предком Anc, имея только одну мутировавшую позицию – вторую с конца. Таким образом, сиквенс Taxon4 – предок сиквенса Taxon3, что, впрочем, не мешает обоим «жить» в одно время, ведь мы сейчас рассматриваем не реальные организмы, а участки их ДНК, которые могут мутировать с разной скоростью, вследствие чего среди живых потомков Taxon4 есть как неизменные, так и мутировавшие до состояния Taxon3. Более того, среди ныне живущих организмов вполне может встретиться и носитель предкового типа Anc, хотя со временем количество таких неизмененных вариантов убывает.
«Итак, мы провели простейшую филогенетическую реконструкцию – восстановили вид общего предка трех сиквенсов, при этом столкнулись с неопределенностью порядка возникновения мутаций и видели, как добавление новых данных может изменить точность нашего анализа» (А.Чубенко, «Сыновья Адама»).
Данный вывод, на мой взгляд, весьма важен.
Обнаружение останков ранее неизвестных видов и подвидов предшественников человека, как мы могли убедиться ранее, способно приводить к весьма существенному пересмотру взглядов палеоантропологов на эволюционное древо человека. В частности поэтому так бурно обсуждаются находки, связанные, например с «денисовским» человеком и «хоббитом». Точно так же (как теперь выясняется) дело обстоит и с результатами генетических исследований.
Ситуация усугубляется тем, что в реальности даже у современного человека считается полностью расшифрованной лишь мтДНК (да и то со второй попытки – в первой позднее была обнаружена ошибка). Для неандертальца и денисовского человека удалось расшифровать ДНК лишь частично, а для более древних останков ДНК выделить пока еще не удается. То есть мы имеем дело как раз с ситуацией, при которой отсутствует масса данных. И появление новых данных, способных повлиять на выводы даже кардинальным образом, очень даже возможно. В собранной же на текущий момент «мозаике» эволюционного древа Гомо сапиенс вполне могут отсутствовать весьма важные детали!..

Рис. 148. В мозаике могут отсутствовать важные детали
Пойдем далее.
«…даже самая близкая эволюционная связь сиквенсов не является непосредственной в физическом смысле слова, так как степень реального родства живых организмов можно восстановить по ДНК только в том случае, если известна скорость мутирования рассматриваемого участка. Встречаются участки ДНК, где мутации определенного типа происходят раз в несколько столетий, и наоборот, бывают крайне консервативные участки, например кодирующие критически важные для организма функции, где большинство мутаций влечет нежизнеспособность нового организма, и поэтому новые «устойчивые» варианты появляются раз в несколько тысячелетий. Специалисты, имеющие практический опыт работы с сиквенсами, даже интуитивно умеют определять примерную степень родства генетических линий, исходя из свойств рассматриваемых участков ДНК, скорости их эволюции и особенностей филогении» (А.Чубенко, «Сыновья Адама»).
Хм… Ничуть не сомневаюсь в профессионализме упомянутых в цитате специалистов. Но интуиция есть интуиция – она может и подвести. И наличие подобного субъективного фактора настораживает. Как теперь разобраться, где специалисты сделали вывод на основе объективных данных, а где – на собственной интуиции?..
Оставим собственные субъективные сомнения в стороне и перейдем к другому примеру, приведенному в статье.
Рассмотренный ранее пример достаточно тривиален и позволяет реконструировать эволюционное древо. Но изменим выборку данных, сохранив первые два сиквенса и добавив два новых:
Taxon1 AGCCTGGATCGTATTAGCATGTA
Taxon2 AGCCTGAATCGTACTAGCATGTA
Taxon5 AGCCTGGATCGTACTAGCATGTA
Taxon6 AGCCTGAATCGTATTAGCATGTA
Изменяющихся позиций осталось вместо четырех всего две – седьмая и четырнадцатая с начала. Однако они имеют неприятную особенность – в них представлены все четыре возможных сочетания изменяющихся нуклеотидов: GT, AC, GC и AT. И какой из четырех вариантов ни выбрать в качестве предкового, придется допустить, что по крайней мере одна из позиций мутировала дважды. Например, если выбран предок GT, от него можно произвести с одной заменой GC, от GC – AC, и далее AT, то есть в четырнадцатой позиции имели место переходы T → C → T. В таких случаях говорят, что имеет место так называемая гомоплазия в паре позиций 7 и 14, причем позиция 14 мутирует «обратно».
Если же в качестве корня выбрать сиквенс с GC, то гомоплазия вызовет уже не обратную, а «параллельную» мутацию в четырнадцатой позиции – в двух ветвях независимо друг от друга происходит мутация C в T: T ← C → T. И чем больше таких пар позиций в выборке, тем сложнее и ненадежнее реконструкция истории таких сиквенсов.
«Заметим, что чем быстрее мутирует позиция, тем чаще она вступает в такие гомоплазийные пары с другими позициями. В реальных выборках гомоплазия встречается достаточно часто, а в случаях, когда рассматриваются участки такого вида, как например тандемные повторы, то почти каждая позиция (или признак) может вступать в отношение гомоплазии с какой-то другой позицией (признаком). Филогенетические программы возвращают результат в виде эволюционного дерева либо множества равновероятных деревьев» (А.Чубенко, «Сыновья Адама»).
Итак оказывается, что возможны случаи, когда генетические исследования дают вовсе не однозначный ответ, а сразу несколько вариантов, вероятность реализации которых вдобавок одинакова!..

Рис. 149. Витязь на распутье (Васнецов)
Далее. Физическая мутация всегда имеет направление изменения – одно состояние цепи ДНК сменяет другое. Эволюционное древо также имеет «корневой узел», с которого началась эволюция рассматриваемой ветви, и «узлы-листья», являющиеся ныне живущими потомками корня.
В первом из рассмотренных выше примеров был использован простейший метод реконструкции предка – в каждой позиции выбирался нуклеотид, представленный у большинства сиквенсов. В том же случае, если мы рассматриваем реальную выборку из большого числа сиквенсов, описанный подход будет неприменим, поскольку трудно предположить, что, скажем, 100 сиквенсов различного вида развились от одного общего предка одновременно и независимо. Скорее наоборот – наверняка в этой выборке есть более близкие друг к другу типы и наоборот более удаленные друг от друга. Следовательно, эволюционное древо может иметь сложную иерархическую структуру.
Вынужденные работать в столь непростых условиях, генетики разработали множество формализованных методов реконструкции структуры дерева, определения вида предковых узлов и корня, которые применимы как к малым выборкам из нескольких сиквенсов, так и к большим выборкам в несколько тысяч образцов. И конечно же, никто не занимается столь сложным анализом «вручную» – для реконструкции дерева используются компьютеры, анализирующие исходные выборки сиквенсов по определенным алгоритмам. И в этих алгоритмах приходится учитывать массу дополнительных деталей.
«…нередко возникает потребность как-то ранжировать позиции по скорости мутаций в них, чтобы программа могла искать деревья, более вероятные с точки зрения известных биологических особенностей мутирования, поэтому предварительная обработка молекулярных данных обязательно включает в себя анализ сиквенсов как последовательностей нуклеотидов. Кроме того, многие методы требуют данных об относительной вероятности перехода одного нуклеотида в другой, притом для каждой позиции отдельно» (А.Чубенко, «Сыновья Адама»).
Вышеприведенные словосочетания «формализованные методы», «использование алгоритмов», «ранжирование позиций», «вероятность перехода одного нуклеотида в другой» подразумевают под собой не только неоднозначность решения (как для последнего словосочетания), но и сильнейшее влияние субъективного фактора – ведь делает это все человек. И методы с алгоритмами разрабатывает, и позиции ранжирует, и даже вероятность перехода оценивает. И использование компьютеров не только не улучшает ситуацию, а наоборот – только усугубляет ее.
Обычный человек часто воспринимает выводы, полученные с использованием компьютеров, чуть ли не за истину, а результаты компьютерного моделирования чуть ли не за достоверно установленный факт. Грешат таким заблуждением, увы, и многие из тех, кто причисляет себя к сообществу ученых.
Компьютер действительно не обладает какими-то субъективными предпочтениями. Но он исполняет то, что ему задал человек – программист, разработчик модели или тот, кто задавал исходный алгоритм для расчетов. Ведь у компьютера нет собственного интеллекта. Что в компьютер заложишь – то и получишь. Все субъективные ошибки человека, попавшие в программу, компьютер в ходе расчетов сохраняет.

Рис. 150. Что в компьютер заложишь – то и получишь
И еще один момент.
При исследовании родословной человека и анализе путей расселения наших предков по разным континентам генетика использует такие термины как гаплотип и гаплогруппа.
«Гаплотипом называют произвольный вариант какого-либо участка ДНК, в контексте, когда на этом участке встречаются вариации нуклеотидов… Гаплотип – понятие эмпирическое, так как последовательность нуклеотидов на каждом участке можно узнать с помощью секвенирования, применяя минимум теоретических усилий.
Набор родственных гаплотипов, происходящих от общего предка, называют гаплогруппой, но это понятие уже не столь элементарно. Очевидно, что огромная неопределенность скрывается в понятии «родственный», ведь… факт филогенетического родства сиквенсов может иметь разную степень достоверности.
Далее, почему не считать, что каждый гаплотип на филогенетическом древе вместе со всеми своими потомками образует гаплогруппу? Действительно, пока не указаны строгие правила выделения «главных» ветвей, такой подход кажется разумным. На первый вопрос (о достоверности гаплогрупп) можно ответить следующим образом: любая гаплогруппная номенклатура основывается на некотором варианте дерева, принимаемом большинством специалистов, поэтому в конечном счете проблема упирается в наличие консенсусного мнения. Например, авторитетным считается мнение авторов Y-хромосомной филогении http://isogg.org или филогении мтДНК http://phylotree.org» (А.Чубенко, «Сыновья Адама»).
Итак, мы вышли на то, что решение о содержании базового понятия и выстраивание эволюционного древа принимается не на основе строгих выводов из объективных фактов, а… методом голосования среди авторитетов (вдобавок, среди авторов самой методики!). К каким ошибкам это может приводить, мы ранее уже рассматривали…
«Молекулярные часы»
Описанная выше методика (при всех ее недостатках) позволяет построить модель эволюционного древа. Но эта модель отражает лишь относительную последовательность и не привязана к абсолютному времени. Для того же, чтобы определить время, когда произошло то или иное разделение ветвей на эволюционном дереве, генетики используют так называемую гипотезу «молекулярных часов».
Гипотеза «молекулярных часов» была выдвинута в 1962 году американскими учеными Эмилем Цукеркандлем и Лайнусом Полингом на основе результатов их исследований структуры таких белков, как гемоглобин и цитохром С. Основная функция гемоглобина – перенос кислорода и углекислого газа, а цитохром С выполняет перенос электронов в дыхательной цепи митохондрий и выполняет еще ряд других функций. Эти белки (как и другие) состоят из аминокислот, выстроенных в определенную последовательность. Сравнивая такие последовательности у разных видов современных животных и людей, Цукеркандль и Полинг пришли к выводу, что количество аминокислотных различий в гемоглобине и цитохроме С растет линейно со временем.
Рис. 151. Цитохром С
Различия в аминокислотных последовательностях появляются в результате мутаций и тесно с ними связаны. И вывод Цукеркандля и Полинга основывался на том, что зависимость между числом мутационных шагов (которые оцениваются по числу различий в аминокислотных последовательностях белков) и временем дивергенции (временем расхождения на эволюционном древе) организмов, из которых эти белки были выделены, является линейной – данные для разных организмов ложатся на прямую линию, что и отражает постоянную скорость процесса мутаций в аминокислотной последовательности.
«Возьмем, например, цитохром С млекопитающих и рептилий. Палеонтологическая летопись показывает, что звероподобные рептилии дивергировали от других рептилий примерно 300·106 лет назад. Цитохромы ныне живущих млекопитающих отличаются от цитохромов ныне живущих рептилий примерно 15 заменами на 100 аминокислот. Следовательно, в этом случае на возникновение 15%-ного различия понадобилось 300·106 лет, или 20·106 – для различий в 1%. Время, необходимое для 1%-ной дивергенции по любому белку, Дикерсон (Dickerson) назвал единицей эволюционного времени (ЕЭВ). Для цитохрома С, следовательно, ЕЭВ равно 20·106 лет. У других белков средние скорости эволюции также постоянны, однако абсолютные скорости эволюции у разных белков различны. В частности, для… гемоглобина ЕЭВ равна 5,8·106 лет, а для фибринопептида – всего 1,1·106 лет» (Р.Рэфф, Т.Кофмен, «Эмбрионы, гены и эволюция»).
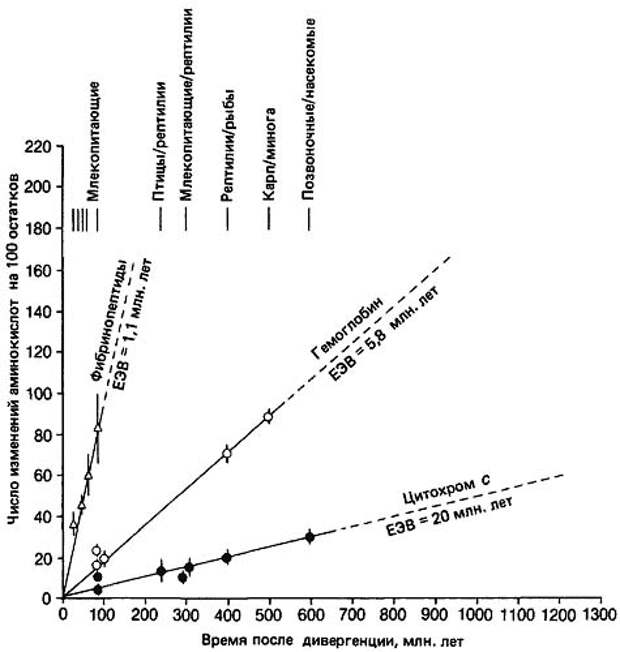
Рис. 152. Постоянство скорости мутаций у различных белков
Отметим, что время дивергенции – количество лет, прошедших с момента, когда произошло разделение двух эволюционных ветвей сравниваемых современных видов (родов и т.п.) – оценивалось по результатам датировок соответствующих палеонтологических находок. К этому моменту мы еще вернемся чуть позже…
Необходимо также сделать следующую важную оговорку. Не следует в данном случае смешивать два разных понятия – скорость мутаций и скорость эволюции. То, что обычный читатель понимает под биологической эволюцией (а профессионалы называют морфологической эволюцией), конечно, непосредственно связано с мутациями, отражающимися в структуре белков, но эта связь имеет сложный характер. Конкретная мутация может иметь разные последствия для эволюционных способностей организма.
Ясно, что если мутация «вредная», то она в конечном итоге приведет просто к вымиранию тех особей, у которых она проявилась. И такая мутация не закрепится в виде соответствующего изменения в структуре белка. «Полезная» мутация приводит к таким изменениям, которые позволяют организму приспособиться (адаптироваться) к изменившимся по неким причинам внешним условиям. Такие мутации называются адаптивными. Но есть и мутации, которые не приносят ни «вреда», ни «пользы». Их называют нейтральными. Количество изменений в структуре белка, возникших в результате именно этих нейтральных мутаций, и является исходными «показаниями молекулярных часов». Некоторые исследователи иногда вводят дополнительно понятие условно нейтральных мутаций, которые, накапливаясь изначально в виде нейтральных, в определенных условиях становятся «вредными» или адаптивными.
«В 1963 году Марголиаш (Margoliash) высказал мысль, что эволюция аминокислотных последовательностей в белках и морфологическая эволюция, возможно, не сопряжены друг с другом. Марголиаш указал, что если истекшее время определяет число замен, накопившихся в данном белке, то эволюция аминокислотной последовательности может служить часами, позволяющими измерить время, прошедшее с момента дивергенции любых двух видов» (Р.Рэфф, Т.Кофмен, «Эмбрионы, гены и эволюция»).
А в конце 60-х годов японский биолог Мотоо Кимура разработал Нейтральную теорию молекулярной эволюции, в которой утверждалось, что подавляющее число мутаций на молекулярном уровне носит нейтральный по отношению к естественному отбору характер. Эта теория также предсказывала существование «молекулярных часов», но при этом не ограничивалась лишь белками, исследованными Цукеркандлем и Полингом, а включала и мутации последовательностей нуклеотидов в ДНК.
Теорию «молекулярных часов» применили в том числе и для определения времени разделения эволюционных ветвей человека и обезьян. Согласно молекулярной оценке, это произошло примерно 5 миллионов лет назад. Такой результат ошеломил палеоантропологов, которые до этого долгое время полагали, что разделение человека и обезьян произошло существенно раньше – около 25 миллионов лет назад. Однако сейчас выводы генетиков все-таки приняты палеоантропологами. Считается, что общий предок человека и обезьян жил 6-7 миллионов лет назад…
Поскольку «молекулярные часы» отсчитывают время по накоплению нейтральных мутаций, не влияющих (по крайней мере напрямую) на видообразование, появляется возможность определять время тех или иных эволюционных событий внутри одного вида – в том числе и Гомо сапиенс. В частности, именно с помощью «молекулярных часов» получены ранее приводившиеся результаты генетиков по миграциям древних людей и освоению ими новых территорий.

Рис. 153. «Молекулярные часы» позволяют заглянуть в прошлое человечества
Нельзя сказать, что теория «молекулярных часов» была всеми сразу принята «на ура». Положенный в ее основу принцип постоянства скорости нейтральных мутаций вызывает серьезные вопросы. Как его можно совместить, скажем, с явно прослеживающимся по палеонтологическим находкам ускорением самого процесса эволюции в целом?.. Или с проявлениями неравномерности такого процесса, как видообразование?.. Так, например, установлено, что скорость видообразования в среднем выше в тропиках, чем в умеренных и холодных широтах (хотя есть и исключения из этого правила) – причины этого давно исследуются, но до конца так и не выяснены. В рамках же теории «прерывистого равновесия» (см. ранее) о таком понятии, как «средняя скорость видообразования», вообще говорить не приходится – она то близка к нулю (на стадии «стазиса»), то становится чуть ли не огромной (на стадии «гнозиса»). А ведь все эти процессы неразрывно связаны с возникновением тех или иных мутаций.
По всем логическим соображениям, в основе возникновения всех («вредных», адаптивных и нейтральных) мутаций должен лежать какой-то единый механизм. Теория же «молекулярных часов» требует принятия идеи, что этот единый механизм приводит сразу к двум принципиально разным процессам – процессу появления «вредных» и адаптивных мутаций, который зависит от целого ряда внутренних и внешних факторов и идет явно неравномерно, и процессу появления нейтральных мутаций, который практически ни от чего не зависит и идет с постоянной скоростью. Такая идея выглядит, как минимум, довольно странной и сомнительной. Особенно с учетом того уже упоминавшегося факта, что нейтральные мутации в некоторых случаях могут стать «вредными» или адаптивными…
Сомнения эти в значительной степени усиливаются тем фактом , что попытки практического применения постулата о постоянстве хода «молекулярных часов» эволюции для анализа хода филогенеза (то есть биологического развития во времени) организмов встретились с серьезными трудностями. Некоторые из полученных этим методом выводов серьезно противоречили сложившимся в науке представлениям, обоснованным традиционными методами и имеющим (как считается) надежную геологическую и палеонтологическую датировку.
Например, среди грызунов дивергенция родов Крыса и Мышь, по данным палеонтологии, произошла не ранее чем 8-12 миллионов лет назад. По данным же молекулярной биологии, дивергенция этих родов определена в принципиально гораздо более древнем интервале времени – 17,5-67 миллионов лет назад (впрочем, такой огромный разброс датировок – почти 50 миллионов лет, который получен в ходе применения разных методов, сам уже говорит против теории «молекулярных часов»). Аналогичное серьезное расхождение дают исследования, проведенные уже в конце ХХ века. Так, «молекулярные часы» указывают для начала дивергенции многоклеточных время 1,5 миллиарда лет назад, между тем как палеонтологи связывают с появлением многоклеточных так называемый кембрийский взрыв – примерно 540 миллионов лет назад, то есть разница в три раза!.. Дивергенция птиц и лепидозавров (надотряд, включающий в себя в том числе ящериц и змей) по «молекулярным часам» происходит еще триасе, а в палеонтологической летописи они появляются лишь в юре (следующий за триасом период). Ветви млекопитающих, ведущие к приматам, грызунам, насекомоядным и хищным дивергируют по «молекулярным часам» уже 140-90 миллионов лет назад – задолго до массового вымирания динозавров 65 миллионов лет назад, которое, по мнению палеонтологов, освободило экологические ниши для млекопитающих.
Подобное расхождение послужило причиной для появления гипотезы, что нейтральные мутации начинают накапливаться задолго до момента расхождения видов, и лишь в некий момент – при изменении внешних условий или при выходе в новую экологическую нишу – нейтральные признаки становятся адаптивными. Но эта на первый взгляд вполне логичная гипотеза влечет за собой весьма неприятные последствия для теории «молекулярных часов».
Во-первых, изначально скорость хода «молекулярных часов» определялась по данным палеоантропологии – по датировкам вполне конкретных находок. Соответственно, для этих находок подразумевалось совпадение момента начала накопления нейтральных мутаций с моментом дивергенции видов. На этом, собственно, и строилось вычисление скорости хода часов. Тогда почему для других видов мы должны допускать отсутствие этого совпадения, да еще и такое сильное?.. И почему принцип более раннего начала накопления нейтральных мутаций не соблюдался для тех видов, по находкам останков которых проводилась «настройка часов»?.. И на основании каких вообще критериев следует в таком случае отбирать палеонтологические находки для «настройки»?.. Может, следует взять совершенно другие и перенастроить «молекулярные часы»?.. Но тогда «поплывут» все полученные результаты и весьма серьезно.
А во-вторых, если принцип более раннего начала накопления нейтральных мутаций, опережающего видовые изменения, имеет место, то он должен быть универсален. Тогда что именно вообще измеряется «молекулярными часами»?.. И какова в этом случае ценность показаний «молекулярных часов», которые показывают нечто, по сути не связанное с реальным расхождением эволюционных ветвей?.. Ответ очевиден – нулевая, как и ценность показаний обычных сломанных часов, которые два раза в сутки все-таки показывают верное время...

Рис. 154. От сломанных часов проку нет
Вместо гипотезы, приводящей к столь неприятным последствиям, был выбран другой путь – начиная с самых первых исследований еще конца 60-х годов ХХ века, давших расхождение показаний «молекулярных часов» с палеонтологическими датами, получило распространение положение, согласно которому «молекулярные часы» идут по-разному на разных ветвях эволюционного древа. Скорость хода часов начали определять для каждой «ветки». Исследования показали, например, что скорость накопления нейтральных мутаций (то есть скорость хода «молекулярных часов») у грызунов оказалась в 2-4 раза выше, чем у копытных, и в 4-8 раз выше, чем у высших приматов. Работа Хэйка по молекулярной эволюции кукурузы показала, что, по крайней мере у некоторых растений, «молекулярные часы» идут на несколько порядков быстрее, чем это обычно наблюдается у животных. И наоборот, у некоторых групп организмов, например у ряда воробьиных Нового Света, ход этих часов замедлен. Аналогичная ситуация наблюдается и в ближайших к нам ветках эволюционного древа – отмечено замедление скорости молекулярной эволюции у гоминид (человека и человекообразных обезьян) по сравнению с другими видами приматов.
Дальше – больше. Выяснилось, что разные белки в одной и той же «ветке» эволюционируют с разной скоростью. А исследования ДНК человека показали, что даже разные гены (участки одной и той же ДНК) накапливают нейтральные мутации с разной скоростью. Причем различия в темпах эволюции различных генов иногда достигают аж трех порядков (то есть скорости мутаций отличаются в тысячу раз)!..
Более того, сейчас уже высказывается мысль, что скорость накопления нейтральных мутаций также изменяется на разных этапах эволюции. Вывод же Цукеркандля и Полинга о линейной зависимости количества мутаций от времени был лишь следствием очень большой погрешности методов «молекулярных часов», которые оцениваются в 20-30 процентов (и это при условии наличия общей тенденции занижать декларируемые погрешности методов). Эта погрешность просто «смазывает» реально существующее непостоянство скорости мутаций. Так, исследования Гудмена показали, что темп молекулярной эволюции увеличивается после генных дупликаций и во времена так называемой адаптивной радиации (адаптации родственных групп организмов к систематическим нерезким однонаправленным изменениям условий окружающей среды).
И наконец еще одна небольшая цитата.
«Постулат о постоянной скорости мутирования не выдержал проверки. Сейчас уже относительно хорошо изучен взрывной мутагенез и, в частности, транспозиционные взрывы [Герасимова и др., 1985]» (В.Красилов, «Нерешенные проблемы теории эволюции»).

Рис. 155. У каждого своя скорость хода «молекулярных часов»
Можно заметить, что ситуация медленно дрейфует к выводу, что нейтральные мутации в общем-то ничем не отличаются от адаптивных как по своей природе, так и по характеру их накопления. А раз так, то мы опять возвращаемся к тому моменту, что в рамках эволюционной теории «прерывистого равновесия», подкрепляемой экспериментами Шапошникова и других (см. ранее), само понятие равномерной скорости накопления мутаций теряет всякий смысл. То, что мы наблюдаем, исследуя относительно недавние изменения – например, при анализе вопроса распространения гаплогрупп современных людей – относится к стадии «стазиса». И попытки применить вычисляемую для этой стадии скорость накопления мутаций на существенно более длительных периодах времени, в которые попадают и кратковременные стадии «гнозиса» (то есть взрывоподобных эволюционных изменений), неизбежно дадут ошибочный результат.
Довольно показательной в данном случае является статья, опубликованная в сентябре 2012 года в научном журнале Nature Reviews Genetics, посвященном вопросам генетики. В этой статье группа исследователей из Бирмингема и Хьюстона призвала пересмотреть принятое значение скорости «молекулярных часов» для человека, которая была рассчитана по степени различия в геномах человека и человекообразных обезьян. И пересмотреть ее в сторону уменьшения интенсивности мутаций аж в два раза!.. Это значение получено ими на основании результатов целого ряда работ, в которых скорость накопления мутаций определялась прямым методом – для современных людей, связанных родственными узами (пары отец-сын или потомки какого-то человека с известным временем его жизни).
Дело в том, что определение времени существования предков современного человека (начиная с гейдельбергского человека) с помощью «молекулярных часов» с традиционным значением скорости дает результат, который как раз примерно в два раза меньше возраста, полученного при датировании реальных палеоантропологических находок. Так, например, костные останки гейдельбергского человека в Атапуэрке (считающегося прямым предком неандертальца), датируются палеоантропологами возрастом 400-600 тысяч лет, в то время как «молекулярные часы» для момента отделения ранних неандертальцев от ветви, ведущей к современным людям, дают всего 270-435 тысяч лет.
Генетические исследования показали, что предки европейцев и жителей Азии покинули Африку около 60 тысяч лет назад. Эти результаты заставили палеоантропологов сделать вывод, что останки древних сапиенсов возрастом 100 тысяч лет, найденные в Израиле, относятся к участникам тупиковой миграции, а не к представителям глобального исхода сапиенсов из Африки.
А если принять предлагаемое в статье снижение скорости накопления мутаций в два раза и взять скорость мутаций у современных людей, показания «молекулярных часов» будут гораздо лучше соответствовать палеоантропологическим датировкам. Для упомянутого гейдельбергского человека скорректированные показания «молекулярных часов» составят около 600 тысяч лет, а время глобального исхода сапиенсов из Африки отодвинется к рубежу 120 тысяч лет назад, и находки в Израиле можно будет уже относить к участникам этого исхода, а не тупиковой миграции (см. Рис. 156).

Рис. 156. Изменение датировок при корректировке скорости «молекулярных часов»
Однако тут возникает другая проблема. В случае проведения этой корректировки при датировании более ранних этапов эволюции вновь возникают серьезные рассогласования с палеонтологическими данными. Так, при замедлении «молекулярных часов» в два раза время существования общего предка человека и орангутана оказывается на рубеже в 40 миллионов лет – чуть ли время существования последних динозавров. Между тем основная масса находок, относящихся к такому предку, датируется палеонтологами временем всего 12-16 миллионов лет назад.
Прямо как в поговорке: хвост вытащили – нос увяз, нос вытащили – хвост увяз…
Для того, чтобы как-то выйти из этого противоречия, исследовали выдвинули предположение, что скорость мутаций замедлялась на протяжении последних 15 миллионов лет. При этом, правда, не уточняется, как именно происходило это замедление и почему…
Между тем, на мой взгляд, ситуация вполне определенно указывает на то, что справедлива именно теория «прерывистого равновесия». И в указанный период времени эволюция человека прошла через стадию «гнозиса», а возможно, и не через одну. Соответствующий всплеск (или всплески) скорости мутаций на стадии «гнозиса» и создает ныне иллюзию более высокой скорости хода «молекулярных часов» в далеком прошлом.
Хотя может быть и иное объяснение, о котором речь пойдет позднее…
Есть ли выход?..
С учетом всего вышесказанного становится неудивительным, что статьи генетиков даже по внешнему виду зачастую больше похожи на произвольное жонглирование цифрами, нежели на строгое научное исследование. И возникает целый ряд вполне естественных вопросов.
Можно ли в такой ситуации полагаться на выводы генетиков?.. Если можно, то в какой степени – до каких временных рубежей и с какой погрешностью?.. И можно ли как-то уменьшить погрешность и расширить временные рамки корректного применения метода «молекулярных часов»?..
Как и в любом другом эмпирическом исследовании, многое упирается в наличие достоверных и проверяемых исходных данных. И тут попытки генетиков улучшить ситуацию можно разделить на два разных варианта.
Первый вариант – повышение достоверности исходных данных достигается использованием для определения скорости хода «молекулярных часов» результатов секвенирования ДНК современных людей. В этом случае исследователи пытаются напрямую измерить скорость накопления мутаций, например, сравнивая ДНК в парах отец-сын у сотен, а то и тысяч таких пар. Также имеют место исследования родственников – потомков какого-то конкретного человека с известным временем жизни. Подобные исследования получили широкое распространение в конце ХХ и начале нынешнего века.
Казалось бы – достоверней данных не найти. Но тут есть две серьезные проблемы.
Во-первых, человек не живет все время с одними и теми же фиксированными последовательностями нуклеотидов в ДНК – мутации накапливаются по ходу его жизни. Какие-то из мутаций передаются его потомкам при их зачатии, а какие-то он накапливает уже после этой передачи. При этом прижизненное накопление мутаций, как выясняется, является не столь уж малым. Например, в исследовании 2009 года, которое осуществлял коллектив из 16 ученых из Великобритании и Китая под руководством Я Ли Сюэ, проводилось сравнение ДНК двух членов китайской семьи, которая живет в одной и той же деревне уже несколько столетий. Ученые взяли для анализа генетический материал у двух мужчин, чей общий предок, отделенный от современников 13 поколениями, жил в XVIII веке. И из всех обнаруженных ими различий в структуре ДНК исследователи отнесли аж две трети к мутациям, возникшим уже в клетках этих мужчин в результате их жизнедеятельности, и только оставшаяся треть была отнесена в категорию «подлинных» мутаций!..
Ясно, что такая ситуация уже создает благодатную почву для серьезных ошибок – как объективных, так и субъективных. А ошибки в определении числа «подлинных» мутаций выливаются непосредственно в ошибки вычисляемой таким образом скорости хода «молекулярных часов».
А во-вторых, прямое измерение скорости накопления мутаций – вещь, конечно, хорошая, но использование ее значения, измеренного для современных людей, при датировании событий сколь-нибудь серьезно отдаленного прошлого может давать очень большие ошибки. Такой прием у математиков называется экстраполяцией, а экстраполяция («продление» функции за пределы диапазона, где эта функция известна) – вещь очень капризная, и использовать ее надо очень аккуратно. Она применима лишь тогда, когда за пределами известного диапазона функция если и изменяет свое поведение, то не сильно. Для случая же «молекулярных часов» это означает, что скорость их хода на протяжении всего рассматриваемого диапазона времени должна быть постоянной или меняться очень незначительно. И уж тем более не должна иметь каких-то резких скачков, как это допускает теория «прерывистого равновесия».
Рис. 157. Экстраполяция может приводить к большим ошибкам
Второй вариант повышения эффективности работы «молекулярных часов» сводится к другому математическому приему, который называется интерполяцией. В этом случае изначально берется большой диапазон времени, для которого определяется некая средняя скорость накопления мутаций, а в дальнейшем полученное значение скорости используется для датирования событий внутри этого диапазона. Этот метод вполне работоспособен, если скорость хода «молекулярных часов» меняется не слишком сильно во всем диапазоне и не испытывает каких-либо серьезных скачков – интерполяция просто «сглаживает» изменения и скачки. Собственно, именно по этому пути и шли генетики с самого начала применения методов «молекулярных часов».
С одним концом такого большого временного диапазона все понятно – это современность. Здесь есть и обширное поле для исследований, и расшифровать (секвенировать) ДНК нужного вида можно достаточно надежно. А как быть с другим концом диапазона, расположенном в прошлом?.. Какую точку на шкале времени нужно взять для того, чтобы свести ошибки к минимуму?..
Как мы уже видели ранее, исследователи брали разные точки и все равно получали серьезные ошибки. Эти ошибки явно указывают на то, что скорость хода «молекулярных часов» вовсе не постоянна, а следовательно брать одну среднюю скорость нельзя. Но ведь (теоретически) можно взять не только крайние точки диапазона, но и некий набор промежуточных точек, для которых определить свое значение скорости накопления мутаций и таким образом получить некую (интерполирующую) функцию. Пусть она и не окажется в этом случае прямой линией, но зато она будет лучше отражать реальное положение дел.
Но это – теоретически. На практике же все сводится опять к вопросу – какие именно точки брать в прошлом (теперь не только крайнюю, но и промежуточные)?..
В публикациях, посвященных генетическим исследованиям и применению метода «молекулярных часов», часто встречается фраза, что для определения скорости мутаций исследователи используют находки с «достоверно установленными датировками». Именно это и определяет начальную точку временного диапазона. Тогда, если мы возьмемся строить некий график изменений во времени скорости хода «молекулярных часов», нам придется добавлять некие дополнительные промежуточные точки, получаемые опять-таки для некоторых находок «достоверно установленного возраста».
Но ведь у нас нет видеозаписей с указателем хронометража для сколь-нибудь серьезно удаленных в прошлое событий. Возможно, когда-нибудь в отдаленном будущем мы сможем изобрести какой-то хроновизор, который позволял бы заглядывать в прошлое. А ныне все неизбежно упирается в имеющиеся методы датирования, о которых мы уже говорили ранее. Ошибки и погрешности этих методов автоматически переходят в ошибки и погрешности «молекулярных часов», не уменьшаясь, а лишь увеличиваясь. И то, что называется «достоверно установленным», на деле таковым вовсе не является…

Рис. 158. Заглянуть в прошлое
На первый взгляд кажется, что чем дальше в прошлое мы отодвинем начальную точку диапазона – тем лучше. Однако в данном случае это оказывается вовсе не так.
Ранее уже упоминалась гипотеза растущей Земли. В рамках этой гипотезы на протяжении последних 250 миллионов лет размеры нашей планеты увеличивались, и увеличивались достаточно серьезно – в целом чуть более, чем в полтора раза (по радиусу). Соответственно, серьезно менялись и условия на планете. Учет этих изменений приводит к необходимости пересмотра всей геохронологической шкалы, а следовательно и «достоверно установленных» датировок палеонтологических находок. И если для периода существования предков человека, оцениваемого ныне всего в 6-7 миллионов лет, коррекция будет относительно небольшой, то для обитателей Земли, живших гораздо раньше, датировки «поплывут» очень сильно. Их возраст может оказаться в несколько раз меньше заявляемого (см. книгу автора «Сенсационная история Земли»). Автоматически столь же сильно изменится и скорость «молекулярных часов» для точек, соответствующих этим древним обитателям. И если уже сейчас имеются случаи, когда показания «молекулярных часов» для дивергенции древних видов превышают в несколько раз датировки палеонтологов, то суммарно для этих случаев придется корректировать «молекулярные часы» почти на порядок!..
Вдобавок, для древних обитателей нашей планеты получить расшифровку ДНК невозможно, поскольку не сохраняется сама ДНК. Поэтому определение скорости мутаций приходится получать не непосредственным подсчетом накопившихся отличий от древней ДНК, а вычислением по структурам ДНК современных потомков древнего вида, которое вынуждено в этом случае опираться на целый ряд дополнительных предположений и допущений. И чем больше таких дополнительных предположений – тем больше возможные ошибки в определении скорости хода «молекулярных часов».

Рис. 159. Выделить ДНК насекомого, застывшего в янтаре, не удается
Если же не удаляться на сотни миллионов лет назад, а ограничиться лишь эволюционным древом предков человека, то влияние расширения Земли можно (в первом приближении) не учитывать. Ситуация кажется несколько лучше. Но это – тоже только на первый взгляд. Ведь, как уже указывалось ранее, для определения возраста находок, относящихся к древним предкам человека, ныне просто нет методов надежного датирования. Уран-торий-свинцовый метод работает лишь на геологических пластах существенно большего возраста. Калий-аргоновый метод не годится для датировки тех слоев осадочного происхождения, в которых обнаруживается подавляющее большинство костных останков предков человека. Радиоуглеродный метод можно считать хоть сколь-нибудь надежным для находок, возраст которых не превышает всего десятка тысяч лет, так что он не годится даже для определения возраста неандертальцев. А все остальные методы датирования так или иначе опираются на перечисленные выше. В итоге практически для всех кандидатов на роль предка человека мы получаем своеобразную «черную дыру» на графике – достоверных точек для настройки «молекулярных часов» здесь получить невозможно. Впрочем, ту же самую «черную дыру» мы имеем даже для ранних Гомо сапиенс…
В результате из всей «человеческой» эволюционной ветви для сколь-нибудь точной настройки «молекулярных часов» нам доступна лишь та часть этой ветви, которая относится ныне антропологами к периоду расселения сапиенсов по планете. Да и то – только последний отрезок этих событий (с учетом реальных погрешностей радиоуглеродного метода) и лишь с большой натяжкой.
Мне пришлось довольно долго искать, что же именно генетики подразумевают под «достоверно датированными» находками, используемыми для настройки «молекулярных часов» периода расселения сапиенсов. В подавляющем большинстве публикаций этот вопрос авторами обходится стороной. В конце концов все же удалось найти статью, в которой указывалось, что такими находками являются… находки на американских континентах, связываемые прежде всего с культурой кловис. И вот тут у меня рухнули последние остатки оптимизма по отношению к «молекулярным часам».

Рис. 160. Наконечники копий культуры кловис возрастом 12 тысяч лет
Дело в том, что когда мы затрагиваем темы, связанные с историей американских континентов, мы попадаем в зону, для которой характерно самое мощное давление со стороны консервативных догматов академической науки. Здесь имеет место самое сильное давление субъективного фактора и самым жесточайшим образом пресекаются даже малейшие сомнения и попытки внести какие-либо коррективы в принятую академическими кругами официальную картину. Между тем как сама картина заселения американских континентов, так и история местных культур не установлена на основе объективных данных, а… просто прописана учеными (преимущественно из США).
Я – не сторонник «теории заговора», но тут термин «принятые датировки», как выясняется, приходится понимать буквально: они действительно принятые в прямом смысле этого слова. Не установленные, не определенные по каким-то деталям или признакам, а именно принятые!..
В это трудно поверить, но это – факт: называемые ныне даты существования индейских культур и цивилизаций – лишь результат откровенного сговора, а не объективных научных исследований.
Например, на протяжении довольно длительного периода в исторической науке были серьезные разногласия по поводу датировок событий и объектов в Мезоамерике. Но в 40-х годах ХХ века было проведено несколько так называемых «круглых столов», на которых ведущие специалисты по истории региона просто договорились между собой о том, как именно они будут датировать известные им артефакты. Постепенно те, кто не согласился с подобным подходом, по тем или иным причинам сошли со сцены, и возмутителей спокойствия не осталось. И с тех пор все новые находки старательно «укладываются» исключительно в прокрустово ложе принятой схемы.
То же, что не вписывается в «утвержденную» картину, просто скрывается. Показательный пример – судьба коллекции шотландского горного инженера Уильяма Невена, который проводил раскопки в долине Мехико в начале XX века. На глубине 9 метров от поверхности он обнаружил следы древней цивилизации с развитой архитектурой, ремеслами и даже письменностью. За время довольно продолжительных раскопок Невеном было собрано порядка 30 тысяч (!!!) артефактов, из которых более двух с половиной тысяч составляли таблички с неизвестной пиктографической письменностью. И вся эта коллекция после смерти Невена в 1937 году исчезла, по сути, в неизвестном направлении (что-то уехало в Нью-Йорк, что-то, по слухам, оказалось в каком-то частном собрании в Мехико, что-то осело в закромах мексиканских музеев). Причина банально проста – подобная цивилизация абсолютно не вписывалась в принятую историками картину прошлого Мезоамерики. И громадная коллекция просто была фактически ими похоронена (почти в буквальном смысле). Если и упоминается где-то сама коллекция, собранная Невеном, то лишь в так называемых неофициальных и «альтернативных» изданиях…
Вот вам размах умалчивания! Тридцать тысяч артефактов!!! Даже в Национальном музее антропологии и истории в Мехико на обозрение туристов ныне выставлено меньше экспонатов!..
В ходе одной из поездок в Мексику нам довелось побеседовать с одним из местных археологов, который в течение почти двадцати лет пытался отыскать хоть какую-то информацию о коллекции Невена, но так и безуспешно. Максимум, что он получал в ответ на свои расспросы, – это совет лучше не затрагивать данную тему вообще, если он не хочет вылететь из системы официальной науки и поставить крест на своей карьере…

Рис. 161. На полках мексиканских музеев коллекции Невена не найти
Аналогичная ситуация наблюдается и местной антропологии. Здесь доминирует точка зрения, что первые люди появились на северо-восточной окраине Северной Америки всего 16-22 тысячи лет назад, перейдя сюда из Азии по сухопутному мосту между этими континентами, который ранее имелся на месте нынешнего Берингова пролива (эту часть суши, существовавшей в то время, часто называют Берингией). Отсюда, как считается, постепенно смещаясь южнее, люди разошлись по обоим американским континентам. Соответственно, в Южной Америке люди могли появиться существенно позднее времени перехода Берингии.
Самые смелые попытки отодвинуть назад во времени заселение этих двух континентов говорят о возможности наличия некоей «первой волны» переселенцев примерно в период 40-50 тысяч лет назад. Но и эти утверждения воспринимаются в академической среде весьма скептически, а датировки соответствующих находок оспариваются. Еще бы – ведь придется кардинально пересматривать все сложившуюся картину расселения Гомо сапиенс, поскольку «достоверно установленный» возраст изменяется более, чем в два раза.
Чуть меньшее сопротивление встречают попытки сдвинуть время появления людей в Северной Америке не на 20-30, а всего-то на пару тысяч лет, но и тут без жестких и горячих споров не обходится. Впрочем, все эти споры в любом случае ограничиваются рамками лишь единственной версии – американские континенты заселялись Гомо сапиенс. Никаких других представителей рода Гомо тут никогда не было. В этом единодушны почти все ученые. Почти…Но все-таки не все.
Продолжение
http://s30662109275.mirtesen.ru/blog/43832356203/10---GENETI...
Находки за пределами антропологических стереотипов
Свежие комментарии